
Arguably, among the most interesting shifts in evolutionary thinking in the past decades is an increased recognition of the limits on rates, directions, and outcomes of evolution. For instance, classic expectations of the adaptive process, such as trait divergence between environments and higher fitness of local vs. non-local individuals, are often not met in natural populations. Effective local adaptation might be hampered by migration and maladaptive gene flow that drags population away from local fitness peaks. Furthermore, instances of (local) maladaptation appear to be particularly pronounced in the context of abrupt and extensive anthropogenic disturbance such as climate change, habitat fragmentation, or pollution. An understanding of the constraints to evolutionary dynamics on the population level therefore needs to become an important cornerstone of ongoing conservation efforts and biodiversity management plans. I here review a recent empirical study from the Hendry lab that investigates constraints to local adaptation in natural fish populations and reveals some interesting contradictions to classical expectations of local adaptation: the “oily guppies” (Poecilia reticulata) of southern Trinidad – a system in which severe crude-oil pollution has a strong impact on the condition of seemingly locally adapted populations.
I first learned about the “oily guppies” at a conference in Berlin in 2010 where Chris Harrod (currently at Universidad de Antofagasta, Chile) presented some interesting results he obtained from guppies that live in oil-polluted streams in the far south of Trinidad. Chris had looked at isotopic compositions of guppies from strongly polluted streams and found that their stable isotope tissue signatures indicated that these fish not only passively tolerate the crude oil, but also seem to actively incorporate it into their metabolism. This was clearly an indication that these fish do not just occasionally “pass through” polluted areas, but dwell in these habitats permanently and, hence, probably show signs of local adaptation to such a severe stressor. Local adaptation to oil pollution – this sounded like a promising research project, not only to me, but also to Andrew, who introduced me to Chris in the first place, and who became my postdoctoral supervisor for the upcoming years of burdensome fieldwork deep down in southern Trinidad.
As soon as (evolutionary) biologists hear the words “Trinidad” and “guppies”, their minds usually wander off to topics such as sexual selection, predation, and life-history trade-offs. All embedded in beautiful northern mountain-range rain forest nerved with pristine crystal-clear streams that provide the researcher not only with ample data but also with refreshing, idyllic... [vinyl scratch sound] Sorry to interrupt, but our story takes place in the south of Trinidad, where things are quite different. Apart from being a Caribbean island with beaches, forests, and Carnival, Trinidad also happens to be located in the southeast corner of the Caribbean plate as part of the eastern Venezuelan Basin – one of the largest oil provinces in the world. Consequently, the Republic of Trinidad & Tobago has been involved in the petroleum sector for over one hundred years undertaking considerable exploration activities, both offshore and onshore (Fig. 1).

Fig. 1. Impressions from southern Trinidad. (1) Onshore oil rigs and hammer pumps. (2) A tour guide demonstrating local geology. (3) High pressure pump systems at Morne L'Enfer forest reserve. (4) Pipeline running through the forest.
Particularly interesting – at least from an ecotox / microevolutionary perspective – are inland drillings that take part in remote forest regions, such as the Morne L'Enfer Forest reserve, where companies aim to mine late Pliocene / early Pleistocene source rock formations and near-surface reservoirs. Obviously, these activities entail major surface leakage and sediment contamination, leaving many tributaries of the region's main rivers heavily polluted with crude oil. I am talking about levels of contamination that are not only apparent and widespread (Fig. 2), but that also make the surroundings smell like a ramshackle gas station. It was here where Chris Harrod, together with Dawn Phillip (University of the Western Indies, Trinidad), had first described populations of guppies from oil-polluted streams. And it was here where I started out my field work endeavors to study local adaptation in fish that live in habitats where (almost) everybody else has jumped ship.

Fig. 2. Pollution impact at our field sites. (1) Morne River – note the oil-soaked black river banks. (2) Field assistant Laura at work – note what the water does to our butterfly nets. (3) Close up of direct leakage into the stream.
After weeks of on-site exploration and many survey hikes, we settled on two promising field sites: Vance River and Morne River, both running through the Morne L'Enfer reserve, and each one with a polluted and a non-polluted tributary respectively (i.e, four sites in total; Fig. 3). At all four sites we observed plenty of guppies and, opportunely, subsequent population genetic analyses revealed that guppies from different rivers can be considered genetically independent (Fig. 4). Moreover, analyses of morphological divergence among the four study populations revealed that similar habitats harbor similar phenotypes: guppies from independent polluted sites would cluster more closely with each other than with their conspecifics from the clean site in their own river system. This pattern was even more pronounced when we focused the analyses on the cranial region (Fig. 5).

Fig. 3. Location of our field sites and the two rivers where we conducted the transplant experiments: MR = Morne River, VR = Vance River; np = not-polluted, oil = polluted.

Fig. 4. Population genetic differentiation among the four studied populations.

Fig. 5. Phenotypic divergence between populations. Shown are warp scores for geometric morphometrics of the cranial region. Along with the four southern populations (MR & VR), we include a northern population (Paria LP, Pa).
Consequently, with confirmed genetic independence and likely parallel phenotypic divergence between similar habitats, everything looked like the perfect set-up for THE standard approach to test for local adaptation in natural populations: a good old reciprocal transplant experiment. Basically, you take individual fish from each of the four sites and transplant them to all other sites (including one “transplant” within the home site), where you then follow them throughout the experiment to see how they are doing – or, more technically, you measure fitness proxies such as growth rate or survival. If populations are locally adapted, you would expect local individuals to show higher fitness outcomes than non-local individuals from other populations. The classic signature in the data would therefore show crossing reaction lines for the direct performance comparison of locals vs. non-locals (Fig. 6).

Fig. 6. Classic local adaptation: Fitness distribution of populations A (dashed line) and B (solid line) in their respective native habitats vs. an alternative habitat.
We did not find this signature in our data. Instead, what we found looked more as if guppies from the polluted environments were doing poorly in all environments. If anything, this was local maladaptation (Fig. 7). What was going on? Well, when your results contradict the expectations that originally led you to set up the experiments, that's where the interesting science starts. Of course, there could have been hundreds of things that went wrong during sampling, transportation, and processing of the experimental fish – as so often in any experimental set-up, especially in the field. Yet, our experiments clearly revealed differences and variation among populations and sites; they also revealed detrimental effects of oil pollution, and we had sufficient numbers of individuals and replicates for each treatment group to draw statistically robust conclusions. But what conclusion to draw from this? At first sight, guppies from the polluted habitats did not seem particularly stressed or unhealthy, and their large abundances in those rivers would indeed suggest that they are locally adapted. But no evidence in our data... well, we needed more data.

Fig. 7. Performance measures from reciprocal transplant experiments. Instead of crossing fitness curves (Fig. 6), we found that guppies from oil-polluted habitats (Origin.oil) do poorly in both conditions (Environment oil vs. Environment not-polluted) when compared to guppies from not-polluted habitats (Origin.np).
Another field season was arranged to repeat the whole transplant design one more time. This time, instead of focusing exclusively on guppies from the described oil-rich region in the south, our second round of experiments included guppies from two pristine northern streams (Aripo & Paria, LP). While it could well be the case that, in general, guppies from the south had a previous history of oil pollution – even the ones from currently non-polluted sites – for sure the guppies from the north were never exposed to any sort of pollution. If there were any carry-over effects or population history interfering with our results, a comparison to pollution-naïve fish should at least highlight any local adaptation signal in our data. This was not the case. Instead, interestingly, guppies from the pristine northern environments were even, to some extent, doing better in polluted-water conditions than the true “pollution-natives” from the south, and when moving “oily guppies” (especially the Morne River population) to clean water conditions, they also performed worse (Fig. 8). Where we expected local adaptation to oil-pollution, we found more or less the opposite.
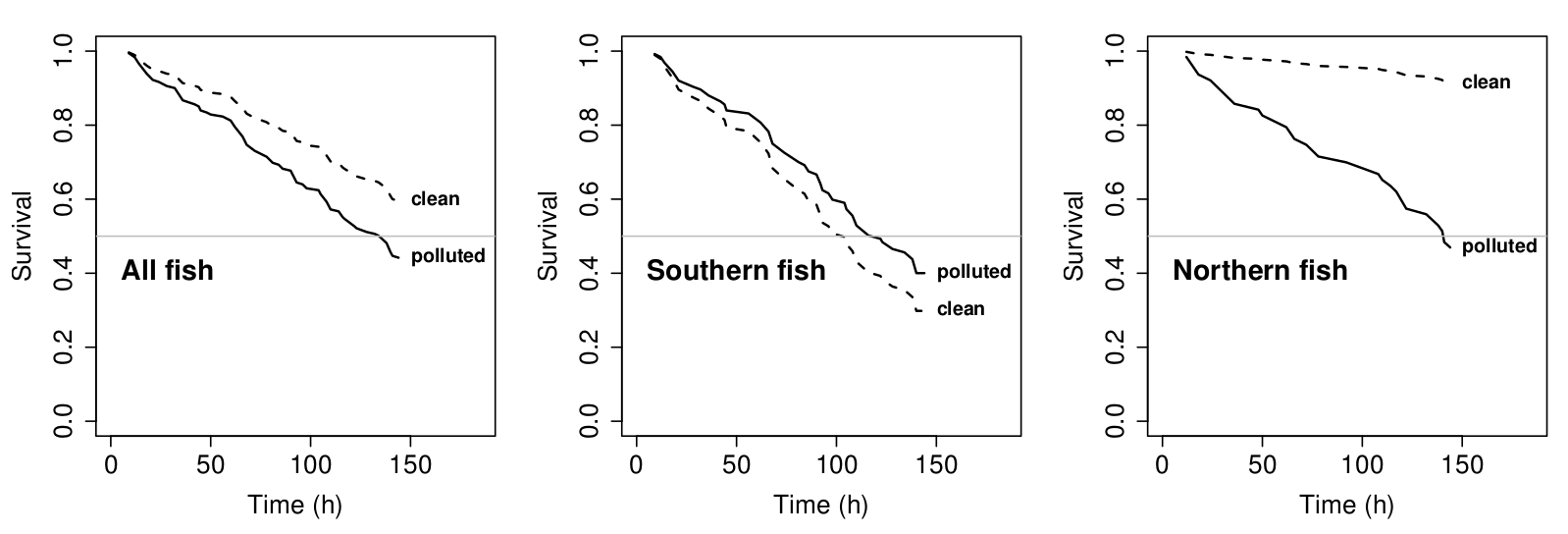
Fig. 8. Survival under laboratory conditions (oil-polluted vs. clean) of guppies from polluted southern habitats vs. guppies from clean northern habitats. (Click to view at full size.)
Now, we have to be careful in speculating about adaptation vs. maladaptation here since we have no data to back up whether the performance differences among our populations have a genetic basis. All fish tested were wild-caught, and thus might have been influenced by plasticity and/or maternal effects. But many examples of adaptive plasticity and maternal effects exist, and given the high selection pressure in our system (mortality), it would still be puzzling to find such an extent of detrimental plastic effects. An even bigger study design would be needed to get to the bottom of this. Another possibility that we did not cover with our study design is that there might be genetic adaptation in early life-stages helping the “oily guppies” to make it to maturity in bad shape (but still functioning), but for the adult phenotypes evolution hasn't come up with a good solution. Moreover, founder effects and/or gene flow from adjacent tributaries might play an important role, but given that our assessment of population genetics and gene flow is currently based on a restricted set of markers, additional analyses with broad-scale genomic tools will have to better inform those hypotheses in the future.
It is, however, equally plausible that crude-oil pollution on the scale present in these rivers is simply too much for the fish to cope with and still be strong and healthy enough to pass the rigorous local adaptation tests. After all, the fish we tested the “oily guppies” against were exposed to oil for a few days only, whereas the pollution-natives face a constant contamination load throughout their lives that might well leave them feeble and vulnerable in their adult life. Interestingly, we did not find many fish species other than guppies in the polluted habitats (Rivulus hartii was there, of course!), nor were there many invertebrates to be found. And indeed, many species are known to have difficulties adapting to highly stressful environments, particularly in the context of anthropogenic pollution and contamination events. Thus, the “oily guppies” could well live on the edge of persistence and, since there is almost no competition or predation, they probably still reach high densities despite poor or sluggish adaptation. Once tested and handled in an experimental set up, the additional stress then pushes them over the aforesaid edge. In this scenario, persistence would not so much depend on the dynamics of adaptation, but more on the fact that the environment is adverse enough to keep away competitors and predators. In addition, we lack information about the exact timing of pollution events. The rivers we studied could have been impacted only very recently, not leaving enough time for the guppies to adapt. Currently, we do not have a final conclusive answer.
Interestingly, around the same time when we were trying to puzzle out why the “oily guppies” from Trinidad were so abundant, yet showed so little evidence of local adaptation, another brave biologist was facing a very similar conundrum: Steve Brady (currently at Dartmouth College) was investigating local adaptation to de-icing salts in amphibians from ponds located near busy roads, where he obtained results pretty similar to ours (you can read his very readable summary of the study at ecoevo-evoeco). Steve's wood frogs from roadside ponds were not only doing worse in non-polluted woodland ponds when compared to woodland populations, they also survived at lower rates in their salt-polluted native habitats – just like the “oily guppies”, they seem to be impacted by the pollutants in a way that causes them to perform poorly in general whenever compared to individuals from undisturbed habitats. Steve, in the search for an appropriate term, called the pattern he found 'deme depression', drawing on the analogous pattern of reduced fitness caused by inbreeding depression.
It goes without saying that our study (as well as Steve's work) on maladaptation is by no means complete, and there are many possible mechanisms that could cause the patterns we find for the “oily guppies”. Yet, I also think that both studies highlight the importance of reporting exceptions to classic predictions, especially in the context of anthropogenic disturbance of habitats. Although negative results, such as the hampered adaptation in “oily guppies“ and wood frogs, are frequently dismissed as resulting from methodological artifacts, it is just as reasonable to think that they actually reflect real biological phenomena that need to be investigated in more detail. We hope that our work encourages further explorations of such exceptions to expected patterns of adaptation in natural populations, especially in the context of stressful anthropogenic disturbances. Understanding the dynamics of adaptation in the wild is of immense practical interest for conservation biology at a time when we are only just beginning to understand our own impacts on evolutionary processes in natural ecosystems. Consequently, our understanding of how and why natural populations sometimes fail to adapt to ongoing environmental changes is an essential target of research that will help to sustain important ecosystem services and inform regulatory decisions. In short, we need not only environmental impact assessments, but also “evolutionary impact assessments” – investigations of the evolutionary consequences of environmental changes on natural populations.
References
Rolshausen, G., D.A.T. Phillip, D.M. Beckles, A. Akbari, S. Ghoshal, P.B. Hamilton, C.R. Tyler, A.G. Scarlett, I. Ramnarine, P. Bentzen, and A.P. Hendry. 2015. Do stressful conditions make adaptation difficult? Guppies in the oil-polluted environments of southern Trinidad. Evolutionary Applications 8: 854–870. DOI: 10.1111/eva.12289.
Brady, S. P. 2013. Microgeographic maladaptation and deme depression in a fragmented landscape. PeerJ. Full text: https://peerj.com/articles/163/




No comments:
Post a Comment