Remember the debate in an earlier blog about whether or not Belgian beers and glasses had co-evolved?
http://ecoevoevoeco.blogspot.com/2011/02/what-did-i-learn-today-lessons-from.html
And the subsequent follow up:
http://ecoevoevoeco.blogspot.com/2011/09/coevolution-of-beer-and-glasses-redux.html
The newest word comes from the champagne world, suggesting that drinks and glasses are not nicely co-evolved:
http://www.themalaysianinsider.com/litee/food/article_food/flute-or-coupe-maximising-the-champagne-experience/
And here is the paper:
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0030628
Thanks to Shahin Muttalib for finding this.

Academic musing by Hendry, Bolnick, Gotanda, and awesome guests. Opinions and statements expressed on this blog are the views of the author(s) and do not necessarily represent endorsement by the blog administrators.
Friday, February 24, 2012
Tuesday, February 21, 2012
The Guppy Project
Today’s post is a quick plug for a great website, The Guppy Project: Experimental studies of the interaction between ecology and evolution in a natural ecosystem. David Reznick writes:
I would add that there are also links to beautiful photographs taken by project members, and a great summary of the scientific motivations behind the project. Enjoy!

Our website summarizes our ongoing experimental studies of guppies, evolution and eco-evo dynamics in Trinidad, W.I. Our project was initially funded by the Frontiers in Integrative Biological Research program (NSF). We are integrating ecosystems ecology, population/evolutionary biology and adaptive dynamics theory. The core project is the experimental study of evolution in natural populations of guppies, with supplementary experiments done in artificial streams and in the laboratory. The website includes a general description of the project, progress reports, PDFs of publications, and an extensive video library. The library consists of videos made on-site in the tropical rainforest of Trinidad. Most videos are two to five minutes long and provide a description of the habitat, natural history and methods of study. There is a series of longer videos that presents an overview of the project, plus a video of a seminar presented by D. Reznick on October 19, 2011 that presents a similar overview and progress. The videos are intended for secondary schools, undergraduates or a general public that wants to know how scientists study evolution in natural populations.
I would add that there are also links to beautiful photographs taken by project members, and a great summary of the scientific motivations behind the project. Enjoy!

Wednesday, February 15, 2012
Eutrophication causes speciation reversal in whitefish adaptive radiations
Following the retreat of the ice after the Würm ice age in Central Europe some 10,000 years ago, Alpine-, and pre-Alpine lakes were colonized by a hybrid swarm of whitefish. These whitefish subsequently sympatrically radiated in parallel into a number of distinct ecotypes in multiple lakes resulting in the existence of between one and six genetically and phenotypically distinct whitefish species within the lakes. Whitefish in general and in this study system in particular have become a key study organism in our studies on adaptive radiation and ecological speciation. Since whitefish are keystone- and dominant planktivores, the system shows great potential for studying eco-evolutionary dynamics and possibly feed-backs.
However, as a result of anthropogenic eutrophication during the last century of almost all larger Alpine and pre-Alpine lakes, whitefish diversity became threatened. In the recent issue of Nature, Pascal Vonlanthen and co-workers show that this eutrophication has led to species loss through speciation reversal. The mechanism responsible for this species reversal is likely limited egg survival in the sediment caused by low oxygen concentration at larger depths as a result of bacterial decomposition of increased organic matter following the eutrophication process. This has differential effects on different species of whitefish, since whitefish diversity to a large extent is maintained by spawning segregation by depth. Diversity loss can happen through two different processes, namely demographic decline and speciation reversal through introgressive hybridization. Vonlanthen and his co-workers show evidence of at least the latter, which is illustrated by decreasing pairwise FST-values between sympatric species pairs. In one specific case, private alleles that were found in a species that went extinct during the eutrophication period are now found in contemporary species.
Eutrophication has especially in industrialized countries been replaced by re-oligotrophication and so also around the Alps of Central Europe. After phosphorous removal in waste water treatment became more efficient and phosphorous in washing detergents was banned, at least in Switzerland
An important question arising from the study is whether we can expect ecosystem dynamics and functioning to return to historic states, when phenotypic diversity has been diminished. Will the trophic transfer rates in the planktonic food web decrease when the most efficient planktivores have been lost? And will utilization of profundal resources disappear or will other species be able to invade these niches before whitefish once again radiate into new distinct phenotypes? But the study also raises interesting questions regarding the process of speciation reversal in it self especially concerning why historically private alleles of species going extinct become introgressed into surviving species. Is the hybridization leading to this process a result of phenotypic plasticity in spawning habitat or time, where previously deep-water spawning species changed their habits and spawned at shallower depths and thereby overlap with other spawning whitefish? Or has a fraction of the distinct species always hybridized, with hybrids historically being selected against due to disruptive selection? In either case, this paper has given us a lot to think about.
The paper:
P. Vonlanthen, D. Bittner, A.G. Hudson, K.A. Young, R. Müller, B. Lundsgaard-Hansen, D. Roy, S. Di Piazza, C.R. Largiader & O. Seehausen (2012). Eutrophication causes species reversal in whitefish adaptive radiation. Nature, 482: 357-362.
Tuesday, February 14, 2012
From the Red Light District to Eco-Evolutionary Stability
I just returned from the Netherlands Annual Ecology (NAEM) meeting, where I spoke about Eco-evolutionary Dynamics, essentially an overview of this emerging field. The meeting itself was in Lunteren, in the central Netherlands, but I spent the first few days in Amsterdam – because the only way to get a reasonably priced flights to Europe is to stay over a Saturday night. In essence, you can either pay for a hotel and meals for a few days or you can pay the same amount to the airline. Not having previously spent any time in Amsterdam, the choice was easy.
 It was damn cold, sometimes well south of -10 C. Now I realize that this isn’t cold compared to Montreal but it sure seemed cold for Amsterdam. Yet the Dutch were loving it. The canals had frozen over and many people were skating on them. Given that the building around the canals were 500 years old, it was almost like some scene out of one of those old Dutch paintings. For someone used to hockey skates, even those straight blades looked like they were a hundred years old. Even if the Dutch were loving it, I got the distinct impression that the birds were not – especially the coots. They were wandering aimlessly over the ice or hunkered down on top – almost as if hoping to melt their way through to the food beneath. As an evolutionary biologist, I immediately thought of that 110 year-old study by Herman Bumpus, where he measured birds that were or were not killed by a severe winter storm in Chicago. That was perhaps the first study to actually quantify natural selection on phenotypic traits in natural populations. I hope someone is out there measuring the coots and other birds that live and die during this event (a postdoc told me how a bunch of his tagged oystercatchers were turning up dead). But the citizens of Amsterdam seemed determined to keep Darwin out of the canals by throwing large amounts of dog food out onto the ice for the coots and gulls and swans.
It was damn cold, sometimes well south of -10 C. Now I realize that this isn’t cold compared to Montreal but it sure seemed cold for Amsterdam. Yet the Dutch were loving it. The canals had frozen over and many people were skating on them. Given that the building around the canals were 500 years old, it was almost like some scene out of one of those old Dutch paintings. For someone used to hockey skates, even those straight blades looked like they were a hundred years old. Even if the Dutch were loving it, I got the distinct impression that the birds were not – especially the coots. They were wandering aimlessly over the ice or hunkered down on top – almost as if hoping to melt their way through to the food beneath. As an evolutionary biologist, I immediately thought of that 110 year-old study by Herman Bumpus, where he measured birds that were or were not killed by a severe winter storm in Chicago. That was perhaps the first study to actually quantify natural selection on phenotypic traits in natural populations. I hope someone is out there measuring the coots and other birds that live and die during this event (a postdoc told me how a bunch of his tagged oystercatchers were turning up dead). But the citizens of Amsterdam seemed determined to keep Darwin out of the canals by throwing large amounts of dog food out onto the ice for the coots and gulls and swans.

Yet even if natural selection was thwarted, sexual selection certainly wasn’t – in the Red Light District at least. As if mocking the weather – and teasing those caught in it – dozens of women in lingerie postured (or sending text messages on their cell phones) from behind glass doors while potential clients shivered and shuffled by outside. Window shopping for warmth! (I would have been more tempted to knock on the door of a kindly old woman selling steaming hot chocolate.) Even church goers could choose (it was Sunday) among dozens of women behind glass doors in a ring of buildings encircling the Old Church; an ever tightening noose of “tolerance” around a last bastion of purity.
But enough of natural and sexual selection in chilly Amsterdam – I was in the Netherlands for a scientific meeting! NAEM was attended by about 300 ecologists of all stripes. My task, along with another plenary speaker (Luc De Meester), was to make a case for the ongoing fusion of ecology and evolution: the “newest synthesis” according to Tom Schoener. In essence, I made the case that contemporary (or “rapid”) evolution has an ongoing role in shaping the short- and long-term dynamics of populations, communities, and ecosystems. At the end of my talk, I outlined a series of predictions for when eco-evolutionary dynamics should be important in natural populations. Many of these predictions logically centered around situations where changes in traits are most rapid and therefore most likely to visibly alter ecological processes.
But I am not so sure anymore – maybe stasis and the lack of change also reflects strong, maybe stronger, eco-evolutionary dynamics. The basic idea is that constancy through time in ecological processes (population size, community richness, productivity, or whatever) or traits might be the result of ongoing evolutionary change that enhances stability. Here are several ways in which this might play out.
- It has been estimated (Burt 1995 – Evolution) that fitness in natural populations declines by 1-10% per generation owing to mutation, maladaptive gene flow, environmental degradation, or competition. And yet fitness does not so decline, implying a lot of evolution each generation just to maintain fitness – and presumably thus population size.
- Eo-evolutionary feedbacks (Post and Palkovacs 2009 – PTRSB) might be stabilizing. By this I mean that an evolutionary change in some trait might alter an ecological variable in a way that imposes selection against further trait change: i.e., a negative eco-evo feedback. In this case, selection ends up stabilizing both the trait and the ecological variable. I have no idea how common such feedbacks are and to what extent they are negative versus positive.
- Phenotypic change might influence the same ecological variable through multiple pathways that offset each other. As just one example, the increasing size of individual plants might increase the number of arthropods but also decrease the number of plants (self-thinning), which might decrease the number of arthropods. Perhaps the outcome would be no net change in arthropod abundance despite plant evolution that has both large direct and indirect effects on those same arthropods. A concrete example of these offsetting effects in guppies will soon be published (Bassar et al. 201 – Am. Nat.).
- Changes in ecological conditions might allow phenotypes to remain constant despite evolutionary change. A particularly cool example occurs when organisms have their metabolic requirements provided by an external source, and so no longer need to maintain their metabolic functions. Parasitoid insects, for example, have larvae that live in a lipid-rich medium and so longer have the need to synthesize lipids themselves – and so have lost the ability (Visser et al. 2010 - PNAS). But the phenotype remains the same. Counter-gradient variation might also fall into this general category.
In each of the above cases, and others, phenotypes and/or ecological processes would remain relatively constant but, if one had turned off the evolution, large changes would have occurred. Perhaps, then, the strongest eco-evolutionary dynamics underlie stability rather than change. On the one hand, this might mean that eco-evolutionary dynamics are even more common than we currently imagine. On the other hand, it also suggests they might be harder to study. Either way, we should expand our research focus so that it isn’t restricted to situations where large changes are observed. Although some have argued that this current focus creates a positive bias toward situations of strong eco-evolutionary dynamics, I am here suggesting that a negative bias is also possible.
Friday, February 10, 2012
Adaptation at the margins
The bit of Dr. Donoghue's talk that I want to discuss here had to do with leaf margins: the edges of leaves. Leaf margins can be described as being "toothed," with serrations, or "entire," with smooth margins. This is perhaps something that you have never thought much about, although you've doubtless seen a good many leaves in your life; but it turns out that, contra Fermat, there may be a truly remarkable proof of evolution and adaptation contained in their margins.
Dr. Donoghue presented some work from his own lab involving Vibernum, a genus of ~170 species of woody plants with a wide distribution in the Americas, Eurasia, and parts of Africa and Oceania. Most viburnums are temperate, but the tropical species are sprinkled across the phylogeny, rather than representing one or a few monophyletic clades. This had been interpreted as showing multiple independent origins of adaptation to tropical conditions from a temperate-adapted ancestor. Clement et al. (2011) constructed a new and improved phylogeny that shows that the opposite may be true: the genus may be ancestrally tropical, and then have diversified into temperate zones.
And herein lies the puzzle. In further work based on the new phylogeny of Clement et al. (2011), Schmerler et al. (in prep) cast into sharp relief an interesting pattern: the correspondence between the climate in which a particular Viburnum species lives, and the type of leaf margins it has (Fig. 1). Despite what appears to be a complex evolutionary history with many independent transitions between tropical and temperate regions, the association between climate and leaf margin is strong across the whole phylogeny.

Figure 1. Association between climate and leaf traits in Viburnum.
Taken from Dr. Donoghue's slides, based upon Schmerler et al. (in prep).
This mystery is larger than Viburnum; that genus is just one striking example of the pattern. Indeed, Peppe et al. (2011) show that plants in colder climates generally tend to have toothed leaf margins, while those in warmer climates tend to have entire leaf margins (Fig. 2). This pattern holds worldwide, and the correlation is quite strong. Several reasons for this correlation have been advanced, having to do with carbon uptake, sap flow, water availability, root pressure, and so forth; there are lots of cites to such explanations in Peppe et al. (2011).
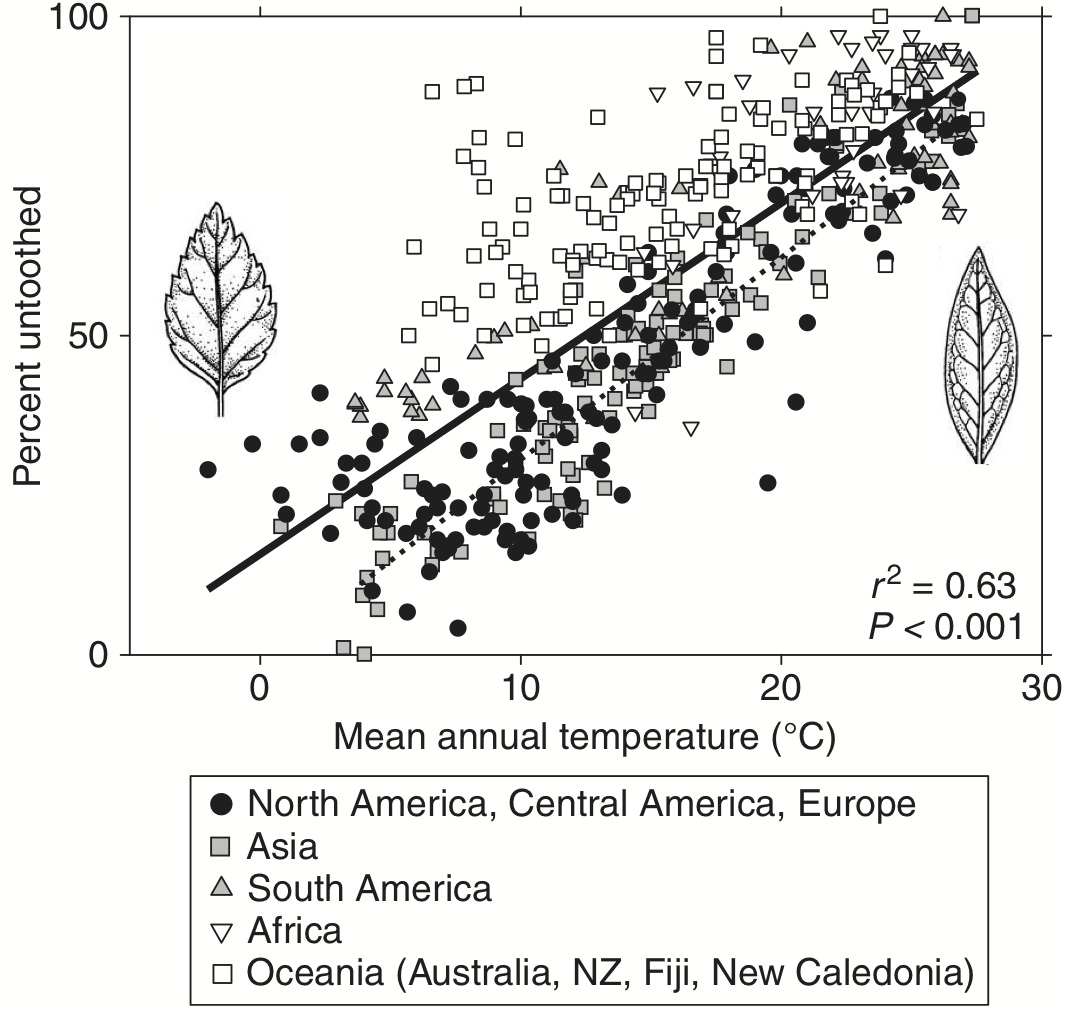
Figure 2. Leaf margin toothedness as a function of mean annual temperature.
Taken from Peppe et al. (2011), their Figure 5; modifications by M. Donoghue.
If I recall correctly, however, Dr. Donoghue said that none of these explanations really seemed terribly convincing. And yet the pattern remained, in need of an explanation. This is where things get really cool.
Couturier et al. (2011) propose that there is a "filling law" that governs the shapes of leaves that grow from buds; in order to pack the developing leaf into the bud as tightly as possible, leaves must have particular shapes, and specifically, their margins must be toothed in particular ways. (Tight packing in the bud may reduce the possibility of frost damage, and the shape and size of the bud may also have effects upon herbivory, if I read them correctly.) They work this hypothesis out in remarkable detail; their paper is a tour de force that essentially founds the new field of biological origami (Fig. 3).

Figure 3. Biological origami: how leaves fold into compact buds.
Taken from Couturier et al. (2011), their Figures 6 and 7.
Their rather fascinating hypothesis, then, is that leaf toothing may have nothing to do with any adaptive value associated with the fully developed leaf; rather, the adaptive value may come from the way that a particular leaf shape folds compactly into a bud. The dynamics of bud growth may determine mature leaf shape, rather than the other way around. And leaves in the tropics may tend to have entire margins because they are evergreen, not deciduous, and do not grow from buds as deciduous leaves do. The colder the climate, the more important it is that leaves are deciduous and begin as tight buds, and thus the more likely it is that the mature leaves will be toothed.
So that's the story. Now, if you will indulge me, I'll give just a few musings of my own.
The reason I like this tale is that it reflects in interesting ways on the idea of the "just so story." Rudyard Kipling wrote his Just So Stories 110 years ago, and they have become a symbol of one of the dangers of evolutionary biology: the "adaptationist" paradigm of assuming that everything must have a reason for its existence — and the subsequent invention, from whole cloth, of said reason. As Stephen Jay Gould and Richard Lewontin (1979) argued so eloquently, traits are not necessarily adaptive. Some things just are, by accident; other things are because they could not be otherwise; and still other things are because their evolution was constrained in some manner — they are because something else is.
In this context, the old explanations for leaf margins might be "just so stories": they might be hand-waving, ad hoc explanations for a phenomenon that was actually, in Gould and Lewontin's terminology, a spandrel. A spandrel is a trait that exists as a result of selection on something else; it is not selected for itself, it is simply carried along. At a genetic level, spandrels might be due to pleiotropy, for example: a gene might have unimportant (non-adaptive) effects on leaf margin, but important (adaptive) effects on some other trait related to climate adaptation, and so adaptation to a temperate climate could produce a correlated change in leaf margins. Such correlations could also be produced by genetic linkage. For this reason, it's dangerous to fall too far into the "adaptationist" or "Panglossian" paradigm of assuming that we inhabit the best of all possible worlds – that every trait is a perfect adaptation to the environment. The old explanations for leaf margins now appear, in hindsight, to fall into this category (at least to some extent; they may also have some truth to them, and I don't mean to prejudge that).
And yet, and yet. What Couturier et al. (2011) have shown us is that even the most obscure, innocuous trait — the serrations at the edges of leaves — may have an adaptationist explanation after all. It isn't the explanation that occurred to biologists at first blush, and so the dangers of the just so story remain very real. But the explanation is there. I don't mean to imply that this is invariably true; I certainly believe that pleiotropy, genetic constraint, sexual selection, and so forth do cause maladaptation in traits, and probably even more commonly cause non-adaptive (neutral) divergence in traits. But the adaptationist program is alive and kicking. Very often, there are reasons for things to be the way they are, and it's worth the effort to look for those reasons. And those reasons, once you finally discover them, are often stunningly beautiful and wholly convincing. Thus it is with leaf margins and the filling law.
As Stephen Colbert might observe, toothiness and truthiness are not so far apart.
References
1. Clement, W.L. and Donoghue, M.J. (2011). Dissolution of Viburnum section Megalotinus (Adoxaceae) of Southeast Asia and its implications for morphological evolution and biogeography. Int. J. Plant Sci. 172, 559-573. doi: 10.1086/658927
2. Schmerler, S., Clement, W., Beaulieu, J., Chatelet, D., Sack, L., Donoghue, M.J. and Edwards, E. (in prep). Evolution of leaf form correlates with tropical-temperate transitions in Viburnum (Adoxaceae).
3. Peppe, D.J., et al. (2011). Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications. New Phytol. 190, 724-739. doi: 10.1111/j.1469-8137.2010.03615.x
5. Gould, S.J. and Lewontin, R.C. (1979). The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist program. Proc. R. Soc. Lond., Ser. B: Biol. Sci. 205, 581-598. doi: 10.1098/rspb.1979.0086
Friday, February 3, 2012
Carnival #44
The 44th Carnival of Evolution is now up at The Atavism. This month's contribution from eco-evo evo-eco is Martin Turcotte's recent post, Why should we care about rapid evolution? (Sorry about it getting attributed to me, Martin; I've emailed them to fix that.) There are lots of other cool posts there, including several on evolutionary ecology and on the timing and tempo of evolution, topics dear to those reading this blog; so check it out!

Subscribe to:
Posts (Atom)
A 25-year quest for the Holy Grail of evolutionary biology
When I started my postdoc in 1998, I think it is safe to say that the Holy Grail (or maybe Rosetta Stone) for many evolutionary biologists w...

-
 As an editor, reviewer, supervisor, committee member, and colleague, I have read countless papers and proposals and have seen similarly co...
As an editor, reviewer, supervisor, committee member, and colleague, I have read countless papers and proposals and have seen similarly co... -
 Bloggy worms? Why worms? Let’s start this post with a small review. Since its start in 2009, the number of contributions on this blog ta...
Bloggy worms? Why worms? Let’s start this post with a small review. Since its start in 2009, the number of contributions on this blog ta... -
 By Dan Bolnick This past month, The American Naturalist published what I hope is the final step in the Editorial Board's evaluation of w...
By Dan Bolnick This past month, The American Naturalist published what I hope is the final step in the Editorial Board's evaluation of w...